
今天跟大家分享一篇文章,来谈一谈实验设计和取样的重要性。这是由香港浸会大学Yiji Xia课题组,发表在the plant journal上的一篇文章。这是一篇典型的基因功能研究的文章,对拟南芥中的一个转录因子bZIP68,在氧化还原应激中的作用机制进行研究。


研究背景和要解决的科学问题
植物抗逆一直是植物学中研究的热点,氧化还原应急则是其中非常重要的一种。细胞氧化还原状态由活性氧(ROS)的产生和清除之间的微妙平衡决定。抗氧化防御系统,包括小分子抗氧化剂和抗氧化酶,在正常生长条件下有效维持细胞氧化还原稳态。胁迫(生物和非生物)可导致ROS的过量产生,从而扰乱氧化还原稳态并对生物分子造成氧化损伤。
生物体已经进化出使用ROS作为信号分子来介导应激反应和发育过程的机制。 ROS信号的传递主要通过富含半胱氨酸的氧化还原敏感蛋白完成的。ROS可以改变这些的氧化还原状态,进而改变这些蛋白的活性,稳定性或亚细胞定位。该课题组前期通过氧化还原蛋白质组学(oxiTRAQ)对拟南芥中的氧化还原敏感蛋白进行鉴定,发现 bZIP68和GBF1可进行可逆氧化修饰。前期的研究也发现bZIP68的突变导致LHCB2的转录水平降低,但bZIP68的生物学功能和bZIP68调控的其他基因仍然未知。
因此,本工作的主要目的就是对bZIP68在氧化还原应激过程中的调控机制,尤其是调控基因表达网络进行全面的刻画。
科学问题拆解
(1)bZIP68是否响应氧化应激?响应的的基团/结构域是什么?
(2)bZIP68在氧化应激条件下,对表型有什么影响?
(3)bZIP68响应氧化应激之后,是通过调控哪些基因影响表型的?
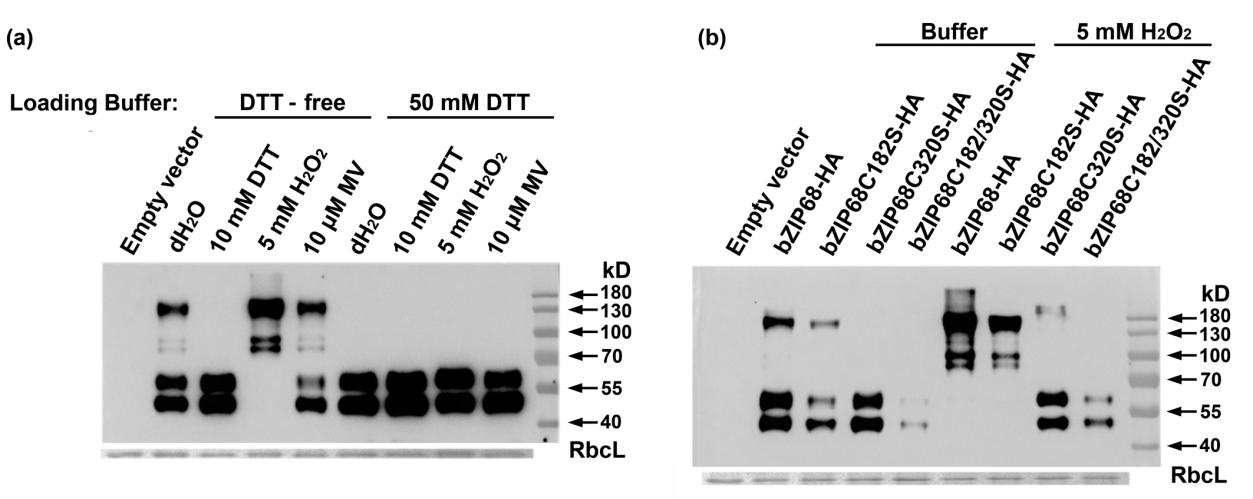
为了回答第一个科学问题,作者首先对bZIP68响应ROS的功能基团进行了确认。bZIP68含有两个半胱氨酸残基,Cys182和Cys320,为了鉴定bZIP68是否在体内氧化应激下经历氧化修饰以及哪个半胱氨酸残基对氧化应激敏感,作者分别构建了带有HA标签的野生型bZIP68蛋白和2个半胱氨酸突变为丝氨酸的蛋白,在原生质体中进行表达并使用H2O2或DTT处理。将处理后的原生质体提取蛋白,使用HA抗体进行WB检测。如果bZIP68在H2O2处理下Cys182和Cys320体内氧化,会形成二硫键,形成大的蛋白复合物;在DTT存在的条件下,二硫键被还原,无大的蛋白复合物产生。
下图可见,野生型bZIP68-HA(44kDa)会形成约150kDa的条带,该条带在样品经DTT处理后消失,说明是二硫键介导形成的蛋白复合物。样品经H2O2处理后,150kDa的条带变成主要条带,说明野生型bZIP68-HA在氧化应激下在体内经历了可逆的氧化修饰。
Cys320被突变成C320S后,不能形成150kDa条带,即使使用H2O2处理后也无150kDa条带出现,表明bZIP68的氧化依赖于Cys320残基。C182被突变成C182S后,蛋白质条带模式与野生型相似,使用H2O2处理后,150kDa条带也变成主要条带,说明bZIP68的氧化不依赖C182S。但结果表明对蛋白的稳定性有一定的作用。
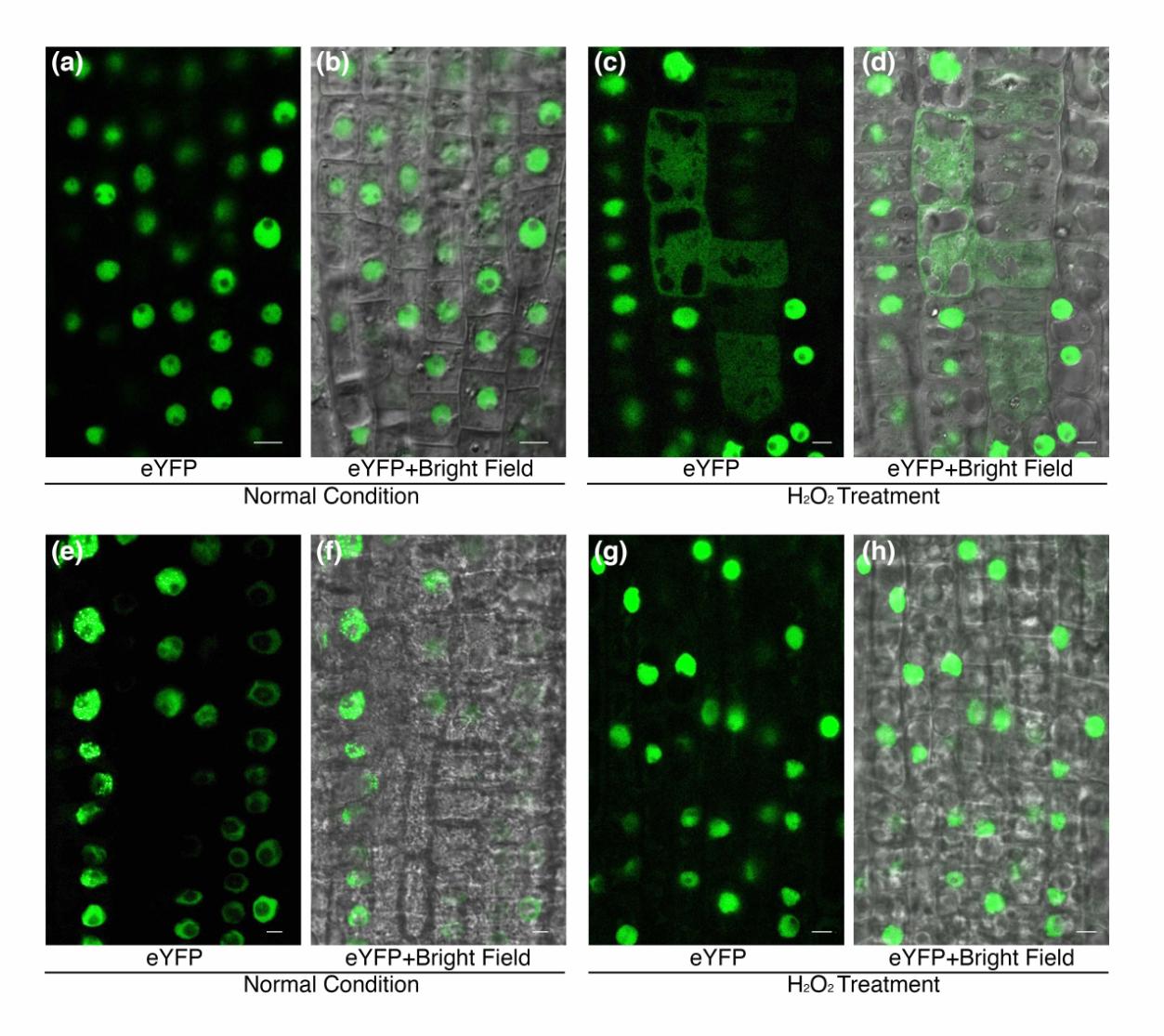
进而作者对bZIP68在氧化应激过程中的细胞定位,以及Cys320对定位的影响进行了研究。通过构建bZIP68和EYFP的融合蛋白并转化拟南芥,作者发现正常生长条件下bZIP68主要定位于的细胞核中;在ROS处理后bZIP68在核中变少,同时出现在根部某些细胞的细胞质中。为了研究bZIP68在氧化应激下的定位是否依赖于Cys320,作者构建了bZIP68C320S-eYFP融合蛋白并进行了转化。与野生型一样,bZIP68C320S-eYFP在正常条件下集中在核内,在氧化应激处理下,bZIP68C320S-eYFP仍然主要留在核内。这表明bZIP68在氧化应激条件下的重定位是依赖于C320的。
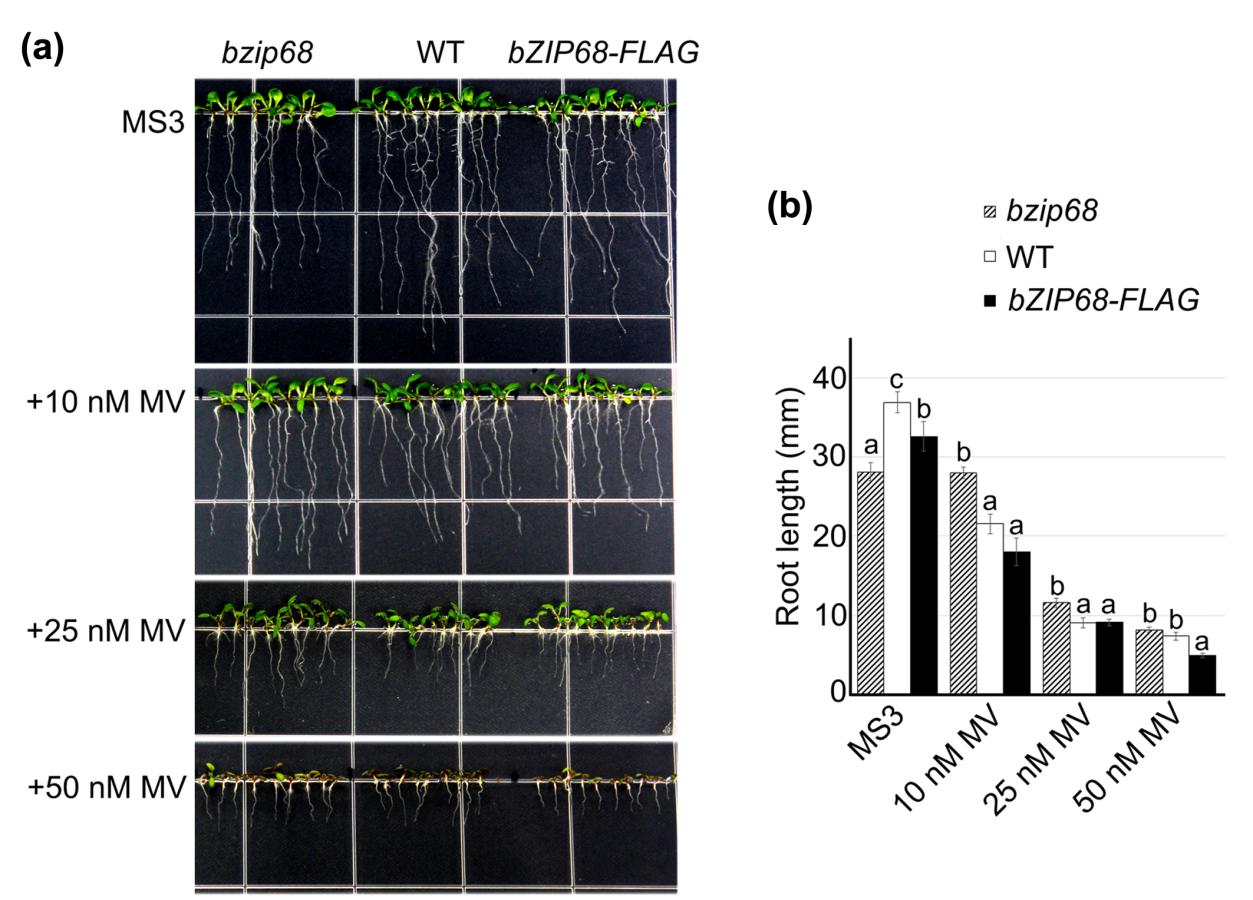
既然bZIP68是响应氧化应激的,那么它是否改变了植株对氧化应激的耐受性呢?对表型的影响是什么?为了回答这个问题作者构建了敲除BZIP68基因的拟南芥突变体株系bzip68,并测试了其在MV存在的条件下幼苗的生长表型(MV,百草枯,可以催化电子从光系统I转移到氧气,导致ROS生产,通常用于诱导氧化应激)。如图所示,虽然 bzip68 幼苗的生长速度比野生型幼苗慢,但在 MV 低浓度的情况下,它们比野生型幼苗生长得更好。而过表达bZIP68的35S:bZIP68-FLAG 转基因株系在MV存在时比野生型存在更严重的生长抑制。
上述结果表明bZIP68的确是在体内氧化还原应激过程发挥作用。
那么,bZIP68是通过调控哪些基因/生物学通路发挥功能的呢?因此文章的最后一部分,作者结合了CHIP-seq和RNA-seq,试图鉴定bZIP68直接调控的基因。
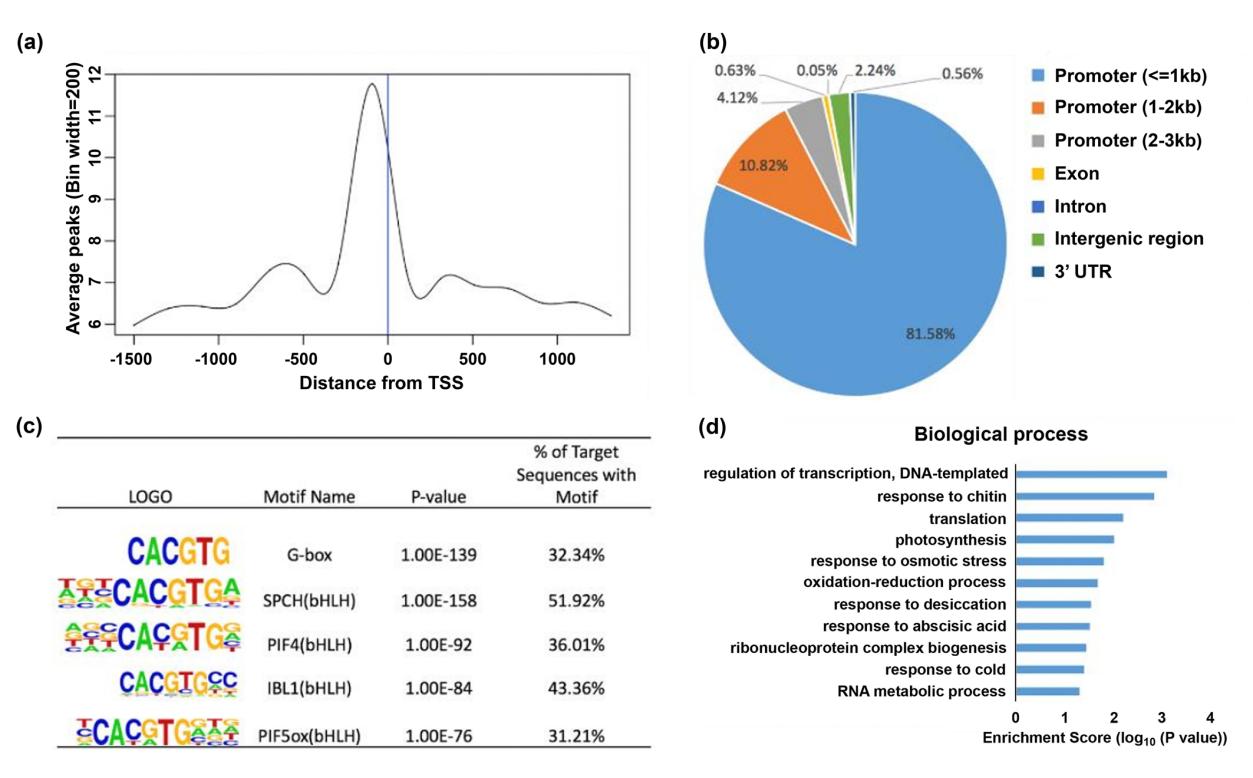
作者在bzip68突变体背景中表达35S:bZIP68-FLAG融合蛋白,使用抗FLAG抗体用ChIP实验,其中包括三个生物学重复和一个Col-0野生型中表达35S:GFP-FLAG作为阴性对照。共鉴定出5,748个bZIP68结合的peak,这些peak与1,058个基因有关。超过80%的peak定位在TSS -1.0 kb至+500bp区间并在TSS的-100 bp 区域高度富集,是典型的转录因子的特征,表明CHIP-seq实验非常成功。进而作者对其中的896 个最强peak所在的 892 个基因进行进一步分析,发现富含G-box and G-box-like motif。该motif是调节生长、发育和对光、stress和植物激素反应的bZIP家族和bHLH家族转录因子的经典motif。对这些基因进行GO分析发现他们主要参与转录调控、对非生物和生物刺激和ABA的反应、光合作用、氧化还原过程,蛋白质/RNA代谢过程等。也包含几个已知的编码抗氧化蛋白相关基因,如过氧化物酶、超氧化物歧化酶和谷胱甘肽过氧化物酶。作者对其中的部分基因/motif使用ChIP-qPCR、EMSA进行了验证。
RNA-seq的数据则表明:对比Mock处理的bzip68和野生型样本,共发现250个基因表达水平存在显著差异,bZIP68突变后,70个基因表达量升高,170个基因表达降低。
WT样本在H2O2处理后,发现 527 个基因表达量升高,189 个基因表达量降低。这些DEG分别被命名为H2O2诱导基因和H2O2抑制基因。H2O2诱导基因中富含heatshock、过氧化氢和其他非生物和生物胁迫应答基因,而H2O2抑制基因富含光、昼夜节律、卡里金、赤霉素,以及细胞分裂相关基因。在突变体中下调的161个基因中,67个为H2O2抑制基因,28种为H2O2诱导基因。以上结果表明,bzip68突变体中的上调基因倾向于H2O2诱导,而下调基因为H2O2抑制。bzip68突变体中H2O2相关基因数目(922)高于野生型数目(716),而且H2O2诱导基因在bzip68中往往被H2O2诱导到比野生型更高的水平,表明bzip68突变增强了H2O2反应基因的表达模式。
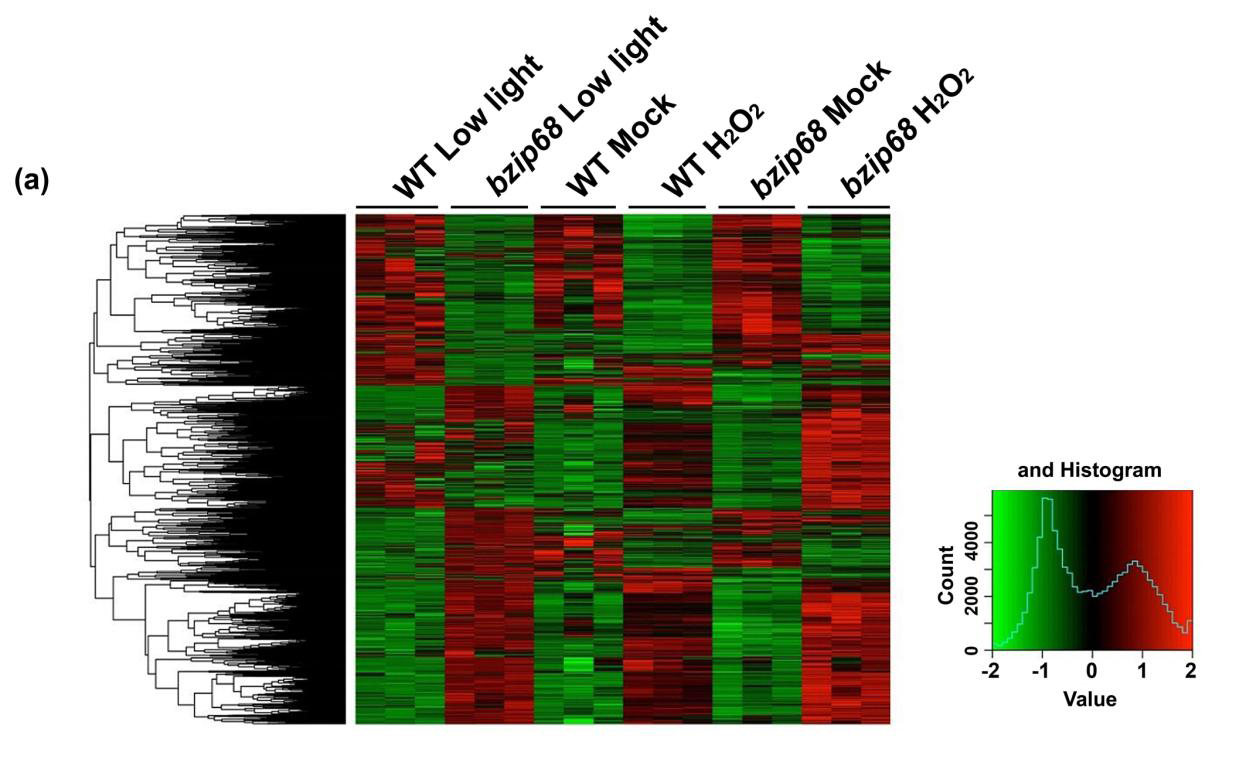
整合CHIP-seq结果和RNA-seq结果,发现ChIP-seq鉴定出的bZIP68结合基因数量大于从RNA-seq鉴定出的bzip68突变型和野生型之间的DEG数:在与bZIP68结合位点相关的892个基因中,只有53个在bZIP68突变后表达发生了差异。暗示bZIP68缺失后大多数差异表达的基因,是受bzip68突变的间接调控。
分析
这篇文章的思路很清晰,工作量也很大,前半部分的工作非常漂亮和可靠,后面的机制研究进行了CHIP-seq和RNA-seq的联用,但是还是有些缺憾:
- 取样问题:
从表型数据可以发现,bzip68基因突变之后,ROS处理表型差异最为显著的是根。但是不管是后面的RNA-seq还是CHIP-seq,使用的都是幼苗。这种取样方式,可能使得突变前后、ROS处理前后根系中有差异的基因,被整株系的基因表达背景湮灭,无法鉴定出来。
- CHIP-seq实验未涉及到ROS
整篇文献只在Mock条件下进行了CHIP-seq,对于ROS处理条件未涉及。因此鉴定出来的是Mock条件下所有可能结合的基因。而对于ROS处理前后,结合发生了什么变化,则无力解析,同时也无法与RNA-seq数据进行整合,这也给后面的分析带来了很大的困扰。
- 研究深度飘忽不定
第一部分作者已经鉴定到了Cys320在氧化还原应激中的关键性作用,但在后续表型、功能研究的时候,直接使用了功能缺失突变体,而作者在CHIP-seq的时候回复表达的也是WT蛋白没有使用Cys320突变。任何一个蛋白的功能都是复杂、多样的,彻底失活基因绝非研究基因功能的最佳手段。
这是一个很好的工作,但如果在实验设计的时候考虑上上述的几个方面,本工作的意义将截然不同。
武汉康测医学检验有限公司 | 专业 ChIP-seq、RIP-seq、MeRIP-seq 服务
地址:武汉东湖新技术开发区高新六路99号武汉市为侨服务产业园项目一期研发楼D2栋1层(1)-6号(自贸区武汉片区)
电话:027-65563640
网站:www.seqmed.cn
邮箱:medical@seqmed.cn

