
研究背景和要解决的科学问题
近日,PNAS上发表了一篇关于研究转录因子的论文,相信对于正在研究转录因子功能的朋友有一定的借鉴意义。
NAC是植物特有的最大的转录因子家族之一,可以通过植物激素信号通路,广泛调控植物的生长发育,并响应多种非生物和生物胁迫。在此之前,从未有人发现NAC转录因子在高温胁迫中的作用,以及干旱和高温胁迫之间的遗传关联。在这篇文章中,作者鉴定了玉米中一个可以介导干旱和高温胁迫的NAC转录因子necrotic upper tips1 (Nut1)。
下面就请大家一起来看看作者是怎么展开研究的吧!

整体思路:
- 研究对象:从突变体库中发现表型变化,找到引起该表型的基因Nut1;
- 功能验证:生物学实验进一步确认突变体引起的表型特征;
- 分析其表达模式,同时为DAP-seq和RNA-seq确定要测序的组织;
- DAP-seq和RNA-seq找到Nut1的靶基因和参与的生物学过程;
- 生物学实验对结果进行验证,证实猜测。
Nut1基因功能
最开始的研究是从作者发现突变体能引起玉米出现干旱表型开始的。作者首先从转座子Activator中分离出隐性nut1-m1突变体。研究发现:
- nut1-m1突变体即使在水分充足的条件下,也出现叶片枯萎、雄穗褐变不育这些典型的干旱和热胁迫表型,并且这种表型在叶尖和雄穗尖端更为严重(下图A);
- 突变体能出现正常的预开花,干旱和热胁迫表型在开花后才开始变得明显,说明该基因可能在花期开始诱导表达。突变体库中的nut1-s1234也出现类似表型(下图B);
- 突变体中也可以看到与干旱和水分胁迫相关的次生生长特征,包括株高降低和穗长减少(下图E)。
维管系统在植物体内可以运输水分,因此作者将与野生型和nut1突变体的维管束进行了比较。突变体维管束的数量虽然减少,但是密度是增加的,说明维管束减少并非是造成干旱和热胁迫表型的直接原因。
那么作者就想,是不是因为突变体水运输障碍才出现这些表型呢?因此作者通过染料示踪实验对此进行了验证:
- 和预期一致,与野生型相比nut1突变体染料运输能力明显受阻(下图C,D),说明nut1基因与水运输相关。

Nut1的组织特异性表达模式
既然已经知道了Nut1与水运输相关,那么Nut1是在玉米的哪些组织、哪些生长时期发挥功能的呢?因此作者使用qRT-PCR对Nut1表达的组织特异性进行检测:
- Nut1在维管组织中高表达,但是在叶尖或雄穗中无表达(图2 B);
- 突变体nut1-m1中未检测到nut1的表达;另一个突变体nut1-s1234表达水平与野生型相当,但没有产生功能蛋白(图2C);
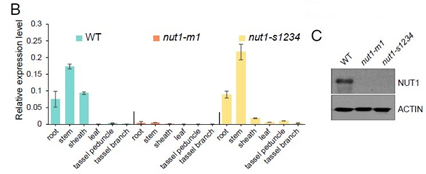
从上述结果也可以看出,Nut1在出现干旱和热胁迫表型的组织部位(叶尖和雄穗)并无表达,说明突变并不是直接影响干旱表型的原因,缺水才是突变表型的直接原因。
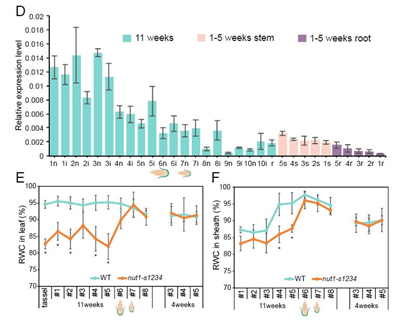
那么nut1在玉米生长周期中的表达情况又是怎样的呢?作者分别对幼龄和11周的玉米不同组织进行检测:
- 在幼龄玉米中高表达Nut1的茎部和根组织连续采样,持续5周,4周后表达量显著增加(图2D),暗示Nut1可能在开花期诱导表达的;
- 作者对数据库中24个玉米干旱实验的RNA-seq数据进行分析,发现Nut1表达无显著差异,证明干旱胁迫处理不能诱导Nut1;
- 在11周龄的开花玉米中,Nut1表达量随茎和节间数逐渐增加,并且在最上面的旗叶(穗下第一片叶子)中高度表达;穗部以上的鞘节和茎节中nut1的表达水平高于穗部以下的鞘节;
- 分别在幼叶和11周龄的中取样,与野生型相比,nut1-s1234突变体的叶片中相对含水量(RWC)显著降低,而在幼叶样本中未见显著差异(图2 E和F);
- 定位分析发现,Nut1蛋白始终在分化为原生木质部的前体细胞核中积累。
以上结果表明,nut1的表达是在原生木质部发育的开花期诱导的,从而增强根茎和鞘中的细胞壁发育并介导水分运输。
Nut1靶基因
Nut1编码NAC91蛋白,属于NAC转录因子家族的成员之一。既然是转录因子,那么nut1能结合哪些基因呢?因此作者使用DAP-seq(和ChIP-seq类似)对nut1的靶基因及识别特征进行鉴定。
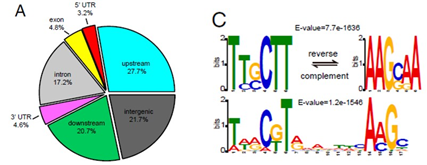
- 共鉴定到5127个peaks,分布于3817个基因之中;大多数peaks (78.3%)富集于基因区域,只有21.7%映射到基因间区域(下图A);
- nut1倾向于与TSS上转录起始位点结合;
- nut1主要识别TTGCTT以及其互补序列,其次是一个17nt的基序。
鉴定到了这么多靶基因,下一步怎么做呢?(记笔记,做ChIP-seq的时候同时做一个RNA-seq)
为了进一步鉴定Nut1的调控网络,作者使用8周龄(此时Nut1转录水平较高,但缺水表型尚未出现)顶端茎和鞘进行RNA-seq,结果显示:
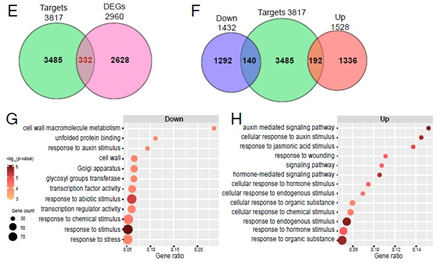
- 与野生型相比,Nut1突变体中发现了2960个差异表达基因(DEGs);与DAP-seq靶基因数据结合,鉴定到332个Nut1的差异表达靶基因(下图 e)。
- 这些基因中包含140个下调基因和192个上调基因(图4F);
- GO分析显示,Nut1下调的差异靶基因主要参与细胞壁大分子代谢,并且与转录因子通路和信号转导相关(下图G);而上调的差异靶基因在激素反应相关通路中富集,特别是生长素和茉莉酸(下图H);
-
这些结果表明NUT1可能多种生物过程促进水的运输。根据以上的结果,作者猜测Nut1可能可以通过控制原生木质部次生细胞壁的生物合成,从而使细胞能够承受在张力下移动水所需的高负压,以此来调控水分运输。
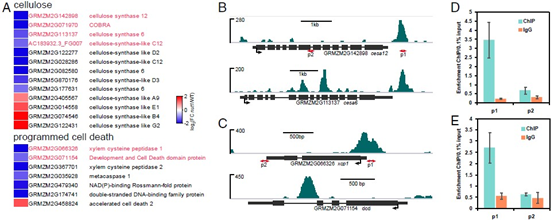
既然Nut1可能是与细胞壁合成相关,而植物细胞壁的主要成分是纤维素和果胶,因此作者进行了下面的研究:
- 对参与纤维素生物合成和凋亡的差异基因进行了分析发现,这些基因大部分都是下调的,其中包含 DAP-seq鉴定到的靶基因(下图A);
- 作者使用ChIP-qPCR对上述靶基因进行验证,证实了Nut1确实能与纤维素合成酶cesa6、cesa12 ,凋亡相关基因xcp1、dcd结合(下图B,C,D,E)。
已有报道显示, cesa12在木质部次生细胞壁生物合成中起作用;xcp1木质部管状分子分化过程中发挥作用,使得植物生成导管,促进水进行长距离运输。
综合上面的结果,说明Nut1能控制纤维素的生物合成,以及通过细胞凋亡因子影响原生木质部的发展。
Nut1功能验证
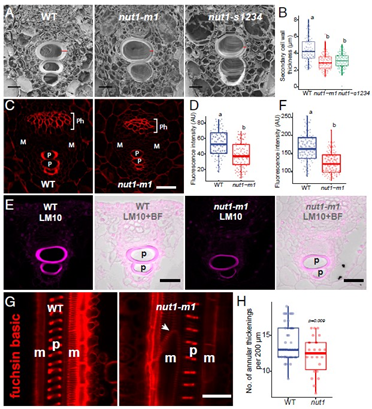
为了进一步验证Nut1能靶向次生细胞壁,参与原生木质部次生细胞壁的生物合成和凋亡。作者通过冷冻切片扫描电子显微镜(SEM)检测原生木质部细胞,发现在nut1突变体中成熟原生木质部导管的次生细胞壁厚度较野生型更薄(下图A,B)。
考虑到Nut1直接控制纤维素合成酶,作者检测了Nut1突变体中是会出现纤维素缺陷。通过纤维素专用染料,比较nut1野生型和突变体纤维素含量。
nut1突变体中的纤维素在原生木质部的次生细胞壁中显著减少,而在次生木质部和韧皮部的纤维素则未受影响(下图C,D)。
使用原生木质部主要结构成分木聚糖的特异性抗体LM10进行免疫定位,发现在突变体中木聚糖含量减少(下图 E,F)。
nut1突变体中原生木质部环状厚壁组织变薄,而且数量减少(下图G和H)。
这些结果表明,nut1中细胞壁成分合成缺陷导致原生木质部次生细胞壁环状厚壁组织异常。
从理论上讲,原生木质部的缺陷并不能完全阻碍整个维管束中的水,次生木质部部应该仍然有运输的能力。因此作者通过透射电镜(TEM)检查了次生木质部的细胞壁,结果发现
- nut1突变体次生木质部细胞壁较薄且表面不平整;
- nut1突变体中原生木质部细胞壁的缺陷导致了中质部的失效和导管连接附近的部分坍塌。
以上结果说明成熟的植物诱导维管束的水分输送可能是紧密耦合的,并依赖于次生木质部和原生木质部之间的相互作用。
本文总结
综合以上结果,该研究表明NUT1转录因子通过调控原生木质部的发育调控水分运输,从而影响玉米的干旱和高温表型,而NUT1表达的时空特异性有利于农作物应对田间水分的不断变化。nut1基因及其相关遗传途径可能是未来提高农作物抗干旱和高温胁迫的重要靶标。
这篇文章的整体思路也是比较简单的,从引起表型变化的基因开始研究,分析其表达模式,同时通过DAP-seq和RNA-seq找到其参与的生物学过程,最后对这个过程进行验证,是一篇比较典型的转录因子功能研究的文章,可以供大家参考一下。

